
近年来,油脂价格居高不下,如何更好地使用油脂成为饲料和养殖企业关注的话题。本文是建明比利时在2021年发表于Animal Feed Science and Technology的综述翻译。希望能够帮助大家全面地了解猪对脂肪的消化和利用。
作者|Alexandra L.Wealleans*,
Karen Bierinckx,
Mauro di Benedetto
译|张智明
摘要
脂肪是猪日粮中主要的能量来源,并且由于成本的增加,高生产性能的基因改良,动物饲料中高品质脂肪的缺乏等原因,营养师对补充脂肪的最大程度利用一直感兴趣。脂肪的消化和吸收是一个复杂的过程,包括乳化、水解及混合胶束形成和吸收三个主要步骤。本文综述了猪对脂肪消化吸收原理的研究现状,以及影响脂肪有效能含量的因素。重要的影响变量包括脂肪酸链的长度及其在甘油三酯上的位置、脂肪饱和度、氧化程度、游离脂肪酸和脂肪含量。这些因素在猪生命周期的不同阶段有不同的影响。本文还对抗氧化剂、外源乳化剂和其他添加剂在猪日粮中提高脂肪利用率的潜力进行了综述。
01
引言
脂肪是一种高能量来源:与脂肪的消化相比,纤维的消化(即使在外源酶的辅助下)释放更低的能量。增加日粮中脂肪的水平将增加猪生长、维持和繁殖所需的有效能量;脂肪的补充降低食糜通过消化道速度,从而有更多时间促进其它营养物质的消化和吸收。然而,脂肪的添加会降低配合饲料的硬度和耐久度。再加上不稳定原料的成本浪费,营养师对最大限度地利用脂肪的能量,从而实现现代猪快速生长和超高产的目标更感兴趣。
可用于饲料生产的脂肪和油种类繁多,包括加工副产品、植物油、酸化皂角和氢化脂肪。脂肪的主要成分是甘油三酯,三个脂肪酸与甘油骨架相连而成。动物饲料中的其它油脂来源组成不同于动植油,如酸化皂角中含有90%的游离脂肪酸。然而,正如Varona(2021)、Wealleans(2021)和Thng(2020)等人所报道的那样,即使是特定来源的脂肪,其组成、氧化状态和适口性等方面也会有很大的不同。此外,众所周知,初级氧化产物的存在会对饲料效率和动物性能产生负面影响。在讨论具有复杂消化过程营养的消化和吸收时,这种成分的变化可能会增加进一步的复杂性。
显然,脂肪的消化过程并不是与肠道中的其它消化过程分开的,而且有多种相互作用影响脂肪的消化率。例如,随着饮食中膳食纤维水平的增加,脂肪的消化率有所下降;这可能是由于饲料中蛋白、脂肪、纤维和其它碳水化合物形成了复合物。相比之下,氨基酸的消化率显示出随脂肪水平的增加而增加。因此,了解脂肪消化和吸收的过程及其影响因素是营养学家非常感兴趣的。尽管脂肪在营养和饲料配方中很重要,但可能由于其来源和组成的多样性,脂肪却成为研究最少的营养成分。本文对影响猪脂肪消化率和吸收率的因素进行了详细的研究,并探讨了提高猪日粮脂肪利用率的潜在策略。
1.1
脂质的分类
脂质分为脂和油。通常,油是指植物来源的脂质,在室温下呈液态,而脂是动物来源,且室温下多为固体的脂质。这些类别也有明显的例外,例如“禽油”。科学上,脂质分为三类:甘油三酯、磷脂和固醇类。甘油三酯是动物营养中脂质的主要成分(约占总重量的98%),次要化合物在总脂肪中所占比例很小。甘油三酯,无论来源如何,都可溶于有机溶剂,主要由碳氢化合物结构组成,即单羧酸。在天然甘油三酯中,酯化脂肪酸有三个立体特异性结合位置,称为sn-1、sn-2和sn-3。
脂肪酸是由它们的碳链长度(C2到C24)来定义的--碳链长度小于6的脂肪酸被认为是短链(SCFA),那些含有6到12个碳的脂肪酸被认为是中链(MCFA),那些超过12个碳的被认为是长链(LCFA)。也可根据它们的不饱和度分类,分为饱和脂肪酸(SFA)、单不饱和脂肪酸(MUFA)和多不饱和脂肪酸(PUFA);有多种命名系统用于描述脂肪酸的化学结构。在营养学中,脂肪酸通常用碳链长度和双键的数量来表示(例如:亚麻酸用C18:3表示)。只有两种脂肪酸--亚油酸和亚麻酸--被认为是猪体内的“必需”脂肪酸(NRC,2012),因为内源合成不能满足生物需求。
短链脂肪酸可内源性产生,是结肠细菌发酵的主要终产物,且浓度较低,通常为58-69 mmol/kg干物质(DM)。若使用富含纤维的饲料可以增加到84-98 mmol/kg DM。短链脂肪酸--尤其是丁酸--为肠道细胞提供了主要的能量来源,并且经常在商业饮食中补充以增加肠道健康。
脂肪酸的碳氢链是非极性的--这种非极性导致了它们的疏水性,如同水是高度极性的,利用溶解分子上的电荷将其他分子留在溶液中。然而,有些脂质因为含有亲水性结构被认为是极性脂,而这种亲水结构使它们至少部分溶于水;这些极性脂是细胞膜的主要成分。从卵磷脂中分离出的极性脂以其乳化能力而闻名,而来自乳制品和动物来源的极性脂是潜在的细胞代谢调节剂。
02
猪对脂质的消化和吸收
猪对脂肪的消化从饲料摄入就开始了,如图1所示。当饲料进入口腔时,它暴露在唾液酶中,唾液酶中包括浓度为2-52 U/l的脂肪酶。在新生仔猪中,唾液和胰脂肪酶活性可能与胃脂肪酶水平相似或更高,但随着猪的年龄增长,唾液脂肪酶的生物学意义逐渐减弱。类似地,Tecles等人(2017)报道了猪唾液中的总酯酶活性水平为75.25-323.65 U/l。总酯酶是一种与脂肪酶类似的酶,专门水解不溶于水的长链甘油三酯。这些酶通过牙齿的研磨作用进一步混合到饲料中,这既从饲料基质中释放甘油三酯,又启动甘油三酯乳化过程。
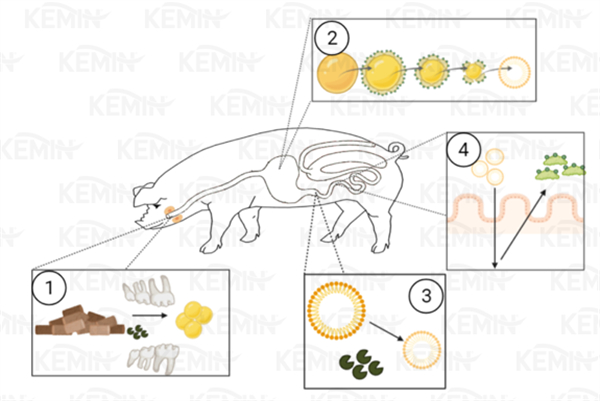
图1:脂质消化三个关键:乳化、水解和吸收的简图概述
1)当饲料被摄入,牙齿的初级研磨将饲料基质中的脂肪释放,唾液脂肪酶被释放。
2)脂肪进入胃后,与游离甘油三酯和脂溶性营养物质混合,形成初步乳状液滴。
3)在十二指肠,进一步乳化,脂肪酶水解甘油三酯为单甘油酯,继而形成混合胶束。这些混合胶束由甘油三酯、游离脂肪酸和脂溶性营养物质组成。混合胶束通过肠壁被吸收,胆盐被释放,最终回到胆囊。
在胃中,甘油三酯主要水解为甘二酯和游离脂肪酸—在此阶段,甘二酯的后续水解可以忽略不计—并且短链游离脂肪酸(FFA)开始直接吸收。这些FFA的释放和吸收是激素级联反应的主要触发因素,包括控制饱腹感和饥饿的CCK反应。在人类中,胃脂肪酶占总脂肪消化的40%(Armand,2007),但猪胃脂肪酶在pH值为4时被不可逆地失活。
尽管唾液和胃脂肪酶都有作用,但大部分摄入的脂肪在猪的小肠中被吸收,进入十二指肠的脂肪有超过70%是以甘油三酯的形式存在。在饲料摄入的一小时内,进入十二指肠水环境中的大多数脂肪仍然没有乳化,而是以大液滴的形式存在。这些脂滴中含有多种脂溶性营养素,如维生素和类胡萝卜素--这些营养素融入脂滴,然后进入胶束结构,这对它们的消化和吸收至关重要。Amiot-Carlin(2009)对脂溶性营养素的消化和吸收进行了详细的综述。
在十二指肠,胆盐和其它双亲化合物(如脂蛋白、磷脂、溶血磷脂)覆盖在脂肪表面,在那里它们开始乳化成更小的脂滴。脂滴进入十二指肠会刺激碳酸氢钠释放到肠腔,并分泌胰酶,包括脂肪辅酶、脂肪酶、磷脂酶和胆固醇酯酶。
在甘油三酯水解发生之前,胰腺脂肪酶必须首先吸附到乳化脂滴的油水界面上。脂肪酶的附着不仅受总有效表面积的影响,还受各种表面活性化合物的影响,这些表面活性化合物包括蛋白质、胆盐、磷脂、单甘酯和游离脂肪酸。由疏水和亲水氨基酸组成的脂肪辅酶,在脂肪酶附着过程中起着关键作用:它能够与脂肪酶形成一对一的复合物,作为脂肪酶的锚点和稳定酶的开放构造,及暴露其活性位点。此外,在缺少脂肪辅酶的情况下,胆盐和其他表面活性化合物会与脂肪酶争夺界面上的有效表面积,从而降低脂质水解的效率。
一旦脂肪酶和脂肪辅酶形成复合物,甘油三酯会被水解成甘二酯和游离脂肪酸。然后甘二脂进一步水解成单甘油酯和游离脂肪酸;这个水解的产物是从sn-1和sn-3位置水解的FFA和sn-2位保留的甘一酯。脂肪酶的活性被其附近积累的甘一酯和FFA所抑制。因此,从界面相中去除这些水解产物是脂肪消化过程中必不可少的一部分。SCFA和MCFA在运输过程中可以作为独立的组分在肠腔内被溶解,而LCFA和甘一酯被转运到水相则需要极性脂质,如胆盐和溶血磷脂。这些极性脂质能够形成混合的胆盐胶束,其中包括甘一酯、游离脂肪酸和脂溶性营养物质。这些混合胶束形成在脂滴界面上的脂肪酶和脂肪辅酶复合物附近。混合胶束是由LCFA、脂溶性维生素和胆固醇酯形成非极性核,然后由甘一酯、游离胆固醇、磷脂和溶血磷脂等极性脂质单层包裹而形成。随后,混合胶束、SCFA和MCFA被运送到小肠的肠细胞。在小鼠身上的研究表明,一些脂溶性营养物质在小肠的吸收比之前认为的更低,趋向于在空肠和回肠远端被吸收,并且——鉴于SCFA和MCFA可被结肠上皮细胞吸收和代谢——这可能与猪是相同的,但还需要进一步的确认。
早期的研究认为,所有脂质通过肠细胞膜的吸收是一个被动的过程,涉及到穿过肠细胞膜的“flip-flop-like”的动作。“flip-flop”运动,也称为磷脂移位,在所有磷脂双分子层中都很常见,并允许膜间磷脂转运。到目前为止,并不是完全了解脂质吸收过程的所有细节(Lairon,2009;Tso等人,2004年),但现在已知SCFA和MCFA是通过简单扩散通过肠细胞膜被吸收的,而LCFA是通过主动的、蛋白质介导的过程吸收的。这种介导的、主动的脂肪酸运输在猪体内的水平没有完全确定。尽管有一个复杂的生理过程,但与其他宏观营养素相比,脂肪相对容易消化。经产母猪的情况尤其如此,因为它们的肠道和微生物区系更加成熟和发达。与生长猪相比,母猪限制饲喂的习惯可以提高脂肪的消化率。
03
脂质在猪日粮中的价值
脂质的有效能量值取决于物种和年龄,并随脂质本身的化学组成而变化。20多年前,一个关于家禽和猪的通用能量预测方程被发表,即广为人知的Wiseman方程。该方程考虑了动物的种类、生命阶段(肉鸡和生长/育肥猪)和日龄(肉鸡为2周和6周,猪的活重为<30 kg和>30 kg)。该方程将家禽的表观代谢能(AME)和猪的消化能(DE)与所用脂肪的饱和度、链长和游离脂肪酸水平联系起来。随后可以根据van Milgen等人的方法计算ME和NE含量。他们提出,对于猪来说,代谢能(ME)是DE的98%,净能(NE)是ME的88%。类似地,Li等人的报告表明,当使用间接量热法测量时,ME是DE的99%,NE是DE的90%。表1描述了Wiseman方程,以及四个常数A、B、C和D对动物种类和年龄的影响。
表1通过Wiseman方程1估算家禽的AME或猪的可消化能DE
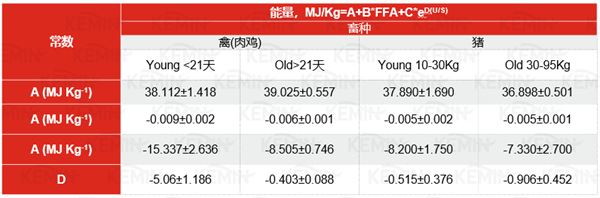
Note:FFA:游离脂肪酸;U:不饱和脂肪酸含量;S:饱和脂肪酸含量;
1改编自Wiseman et al.1998
内源性脂肪损失(ELF)使预测猪日粮中脂质的真实利用率变得更加复杂,ELF是指出现在食糜或粪便中的非膳食来源的脂肪。因此。ELF的估计可以基于单一原料或全价日粮,生长猪的平均ELF值在−1.33—14.02 g/kg DM摄入量之间,不同研究的结果不一致,如表2所示。不同脂质来源之间的ELF差异可能与脂质的U:S比率有关,高比例的不饱和脂肪酸由于促进后肠微生物对不饱和脂肪酸的氢化而导致粪便排泄的ELF增加。
考虑到这些复杂性,以及脂肪来源和内部的成分变化,预测方程必须被认为是普遍的,并没有被证明适用于所有饮食脂肪来源。例如,Boyd等人报道,实验测定的精选白脂的NE值与NRC(2012)的公布值相差14%。Kellner和Patience报告称,通过纳入特定脂肪酸含量可以提高方程的准确性,这表明脂质成分的自然变化会对有效能产生显著影响。此外,工业过程如提取、精炼和裂解会影响化学结构,因此会引起ME和氧化敏感性的变化。此外,由于加工、运输、储存和操作不理想,必须考虑诸如水分、杂质和不皂化物(MIU,例如蜡和复杂脂质)和不可洗脱物质(NEM)等对能量的稀释。
除了提供能量,膳食脂肪也是脂肪酸的来源,包括必需脂肪酸亚油酸(n-6)和亚麻酸(n-3)。脂肪酸组成--包括n-3脂肪酸与n-6脂肪酸的比例--决定了脂质的许多物理性质,经常用于区别不同脂肪,如表3所示。改变猪日粮中n-3:n-6的比例可能显著的影响生产性能:高n-6日粮已被证明会导致母猪生产性能下降,尽管在仔猪身上的证据有限,而且往往相互矛盾,还需要进一步研究不同n-3:n-6比例在实际饲料配方中的影响。
考虑到这些复杂性,以及脂肪来源之间和内部的成分变化,预测方程必须被认为是普遍的,并没有被证明适用于所有饮食脂肪来源。例如,Boyd等人报道,实验测定的精选白脂的NE值与NRC(2012)的公布值相差14%。Kellner和Patience报告称,通过纳入特定脂肪酸含量可以提高方程的准确性,这表明脂质成分的自然变化会对有效能产生显著影响。此外,工业过程如提取、精炼和裂解会影响化学结构,因此会引起ME和氧化敏感性的变化。此外,由于加工、运输、储存和操作不理想,必须考虑诸如水分、杂质和不皂化物(MIU,例如蜡和复杂脂质)和不可洗脱物质(NEM)等对能量的稀释。
除了提供能量,膳食脂肪也是脂肪酸的来源,包括必需脂肪酸亚油酸(n-6)和亚麻酸(n-3)。脂肪酸组成--包括n-3脂肪酸与n-6脂肪酸的比例--决定了脂质的许多物理性质,经常用于区别不同脂肪,如表3所示。改变猪日粮中n-3:n-6的比例可能显著的影响生产性能:高n-6日粮已被证明会导致母猪生产性能下降,尽管在仔猪身上的证据有限,而且往往相互矛盾,还需要进一步研究不同n-3:n-6比例在实际饲料配方中的影响。
表2多项研究中对提取或完整脂质在猪上内源损失的评估
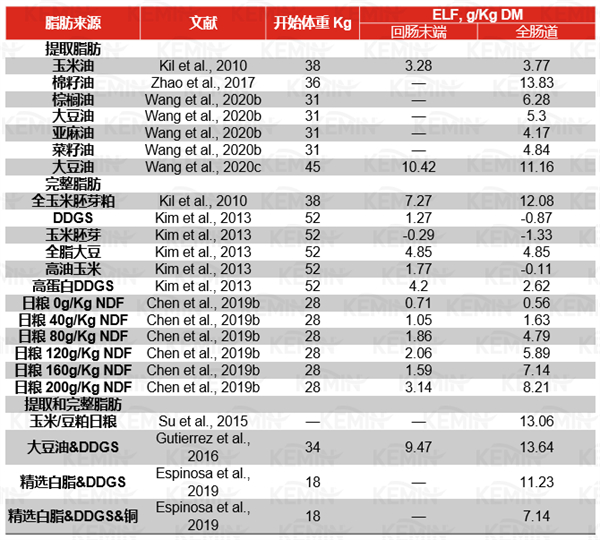
表3非反刍动物常用油脂的脂肪酸分布、PV、TBA和MIU、FFA、U/S等DE计算指标的平均值1
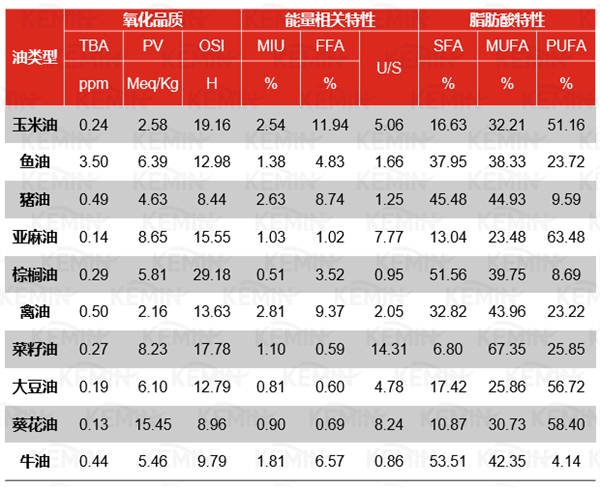
Note:TBA:硫代巴比妥酸值;PV:过氧化值;OSI:氧化稳定性指数;MIU:水分、杂质和不皂化物含量;U/S:不饱和脂肪酸/饱和脂肪酸;SFA:饱和脂肪酸含量;MUFA:单不饱和脂肪;PUFA:多不饱和脂肪酸。
1改编自Wealleans et al.(2021)
最近,Wealleans等人对商业脂质样本的组成和质量进行了分析,发现水分、杂质和不皂化物(MIU)等能量稀释因素的存在会使大豆油、动物脂肪、酸性油、葵花籽油和禽油样品的能值降至公布估算值的46%。这些发现表明,随着优质脂质越来越多地被人类消费,饲料油脂被更便宜的替代品取而代之,猪饲料生产商面临高估脂质能量贡献和质量的风险。由于像MIU这样的成分可能会导致脂质的能量发生相当大的变化,最近发表了一个包含MIU校正因子的修正Wiseman方程,可以更真实地估计脂质的营养价值(公式1)。如前Wealleans等人所述,这一修正因子的加入有时会导致能量变化高达45%。
未完待续……
后续完整内容请持续关注
建明全方位营养!







 警报拉响!养猪人及时做好防控措施
警报拉响!养猪人及时做好防控措施 产能创新低!养户等待好猪价到来时
产能创新低!养户等待好猪价到来时 使用优质保育料,避免掉进这个坑!
使用优质保育料,避免掉进这个坑! 比肩太阳鸟 扬帆饲料展
比肩太阳鸟 扬帆饲料展 对未来有信心!牧原股份:控股股东
对未来有信心!牧原股份:控股股东 脚下有泥,心中有光 | 牧原农艺师
脚下有泥,心中有光 | 牧原农艺师 携手共赢,友谊长存:江西正邦作物保
携手共赢,友谊长存:江西正邦作物保 国新办举行7月份国民经济运行情况新
国新办举行7月份国民经济运行情况新 农业农村部就2022年“三夏”生产形势
农业农村部就2022年“三夏”生产形势